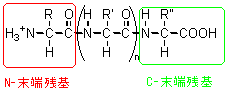Chem-Station-WhatsNew!-BBS-GuestBook-Chat-オークション-サイトマップ
|
|
Chem-Station-WhatsNew!-BBS-GuestBook-Chat-オークション-サイトマップ |
|
アミノ酸(ペプチド)分析 |
|
アミノ酸をつないでいる-CO-NH-結合は、ペプチド結合と呼ぶ。このように結合しているアミノ酸のことをアミノ酸残基という。タンパク質というのはペプチド鎖を持つ分子のことである。
▼ 末端残基
ペプチド鎖は、片方の末端に-NH3+基を持ち、もう一方の末端には-COOH基を持っている。これらをそれぞれ、N-末端残基、C-末端残基と呼んでいる(図1)。
図1 N-末端残基とC-末端残基
▼ アミノ酸組成の分析
まずは、アミノ酸の組成分析について述べる。タンパク質やポリペプチドは6M塩酸を加え、24時間還流すると、ペプチド結合が加水分解され、アミノ酸の混合物が得られる。次に個々のアミノ酸を分離、同定する。アミノ酸は22種類もあるため、これでは手におえない作業となってしまう。
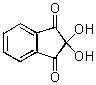
1950年にRockefeller研究所からアミノ酸自動分析装置が開発された。この装置は陽イオン交換樹脂である、不溶性重合体を用いている。アミノ酸混合物をふくむ溶液をカラムに通すと、負に帯電している陽イオン交換樹脂が正に帯電したアミノ酸を吸着する。アミノ酸の塩基性によって吸着力が異なり、塩基性が強いほど吸着力も強くなる。続いて、ある一定のpHの緩衝溶液をカラムに通すと、個々のアミノ酸がそれぞれ異なる速度で降下するので、これによってアミノ酸を分離する。カラムの下端でニンヒドリン(図2)を混合すると、アミノ酸と反応し、強い紫色を呈する誘導体を生じる。アミノ酸分析装置は溶離液のの吸収を連続して測定できるので、その吸収を溶離液の容積の関数として記録できるように設計されている。
図2 Ninhydrin
▼ 分子量、分子式の決定
アミノ酸の組成が決まると、次に、分子量を決定することになる。分子量は超遠心法、光散乱法、浸透圧法など、多種多様な方法が用いられる。 分子量とアミノ酸組成が決定すると、たんぱく質の分子式を決定することができる。たんぱく質中のアミノ酸残基がいくつずつあるかを知ることができる。分子式が決定したからといってこの時点では構造式を決定することはできない。例えば3種類のアミノ酸がひとつずつ含まれているトリペプチドにも3×3×1=6種類存在する。20種類10個のアミノ酸からなるタンパク質においては20100=1.27×10130種類にもなる。
このようにアミノ酸の結合順序の決定を可能にするいくつかの方法が開発されている。このうち、「末端アミノ酸分析」「酵素による分析」を紹介する。
▼ 分子構造の決定
(訂正: 図3、4のペプチドの構造が間違っています。-C(=O)ONH-になっていますが正しくはC(=O)NH-です。)
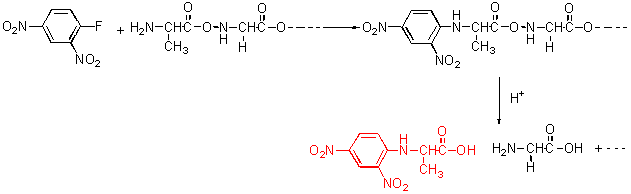
■末端アミノ酸分析 ▼Sanger法 N-末端アミノ酸残基の決定のひとつにSanger法がある。温和な塩基性溶液中でポリペプチドをDNFB(2,4-Dinitrofluorobenzene)を用いて処理すると、N-末端残基の遊離のアミノ酸との間で求核置換反応が起こる。
図3 ポリペプチドとDNFBの反応
その後、酸を用いて加水分解すると、アミノ酸の混合物が得られるがN-末端残基は2,4-dinitrophenyl基が結合している。よって、これを単離し同定することができる。
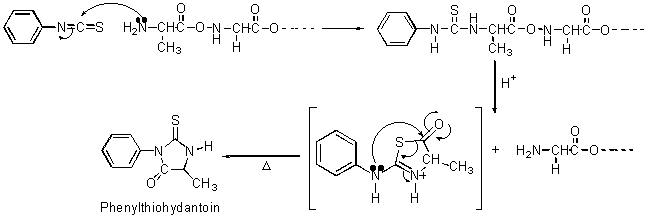
▼Edman分解 次に、Edman分解について述べる。この方法はN-末端残基を取り外した後、残りのペプチド鎖がそのまま残るため、Sanger法よりも有利である。この方法は、N-末端アミノ基とPhenyl isothiocyanateとの標識反応を用いる。標識されたポリペプチドを酸で処理すると、N-末端アミノ酸残基だけが取り外される(図4)。
図4 Edman分解法
得られたPhenylthiohydantoinと標準アミノ酸から合成されたPhenylthiohydantoinを比較してN-末端アミノ酸を同定する。残ったポリペプチドに対してもう一度、分解反応を行うこともできる。この反応は自動化もされている。しかし、N-末端アミノ酸残基を取り除く時に行う酸処理の際に加水分解され生じたアミノ酸が反応混合物中に堆積され、この反応を妨害するため、無限に反復することはできない。アミノ酸残基60個からなるポリペプチドまでは、配列決定装置(Edman法を用いた装置)によって分析されている。
■酵素による分析 Edman分解によって構造決定ができないタンパク質などは、タンパク質を適当な長さに加水分解して、それぞれの構造決定を行った後につなぎ合わせる。ポリペプチドやタンパク質を小さなペプチドに分解するにはいくつかの方法があるが、プロテアーゼというペプチド鎖を選択的に加水分解する酵素がいくつかある。
▼セリンプロテアーゼ この酵素は、活性部位にセリンがあり分解して生成したアシル基がセリンのヒドロキシル基と結合する。この種類の1つにトリプシンがある。ペプチド結合のカルボニル基がリシンやアルギニン単位の一部である時に開裂を起こす。また、キモトリプシンはフェニルアラニン、トリプトファン、チロシンのような疎水性の残基のカルボニル基で形成されたペプチド結合を開裂する。
▼カルボキシプロテアーゼ ペプシン(胃に存在する)はpH1.0で最大活性を示し、2個のアスパラギン酸残基のカルボキシル基を活性部位の官能基として持っている。また、レンニン(チーズ製造に用いられている)も同じ性質を示す。これらの酵素はセリンプロテアーゼよりも特異性が低く、一般に疎水性の残基の間の加水分解を起こす。
▼チオールプロテアーゼ チオールプロテアーゼは活性部位のチオール基がアシル化される。papain(パパイヤ果実に含まれる)、ficin(無花果に含まれる)、bromelain(パイナップルに含まれる)、actinidin(キウィに含まれる)などがある。papainは切断される結合のN-末端側に疎水性アミノ酸残基を持っているようなポリペプチドを加水分解する。
▼臭化シアン 臭化シアンBrCNは酵素ではないが、メチオニン単位のカルボニル基のところでのみ鎖を開裂し、メチオニンはC末端ホモセリンラクトン単位に変換される。
これらの酵素で分解されたポリペプチドをEdman分解によって分析し、それらをつなぎ合わせることによってタンパク質やポリペプチドの構造を決定する。
アミノ酸の構造決定は、単純な作業の繰り返しなので大変な作業らしい。が、やっぱり有機って面白いよね!! (2000/10/13 ボンビコール)
▼参考、関連文献
・ソロモンの有機化学 下 第4版 廣川書店 ・ストライトウィーザー有機化学解説2 廣川書店 ・天然有機化合物の構造解析~機器分析による構造決定法~ シュプリンガー・フェアラーク東京
▼関連リンク
|